





在新时代对基因编辑作物商业化的展望与思考(上)中,笔者谈及了基因编辑的基本概念及其在作物改良上相对于转基因的优势。
那么基因编辑在作物品种改良上能起到多大的作用,哪些性状可以使用基因编辑技术进行改良呢?
虽然我国《农业用基因编辑植物安全评价指南(试行)》刚发布,但作物基因编辑实验早已广泛开展并获得了丰硕的成果,其中的一些代表性成果分类和汇总如下。
一、作物抗逆性改良
自从分子育种技术发展以来,抗逆性改良就一直是最快走向商业化应用的方向。比如在分子标记辅助选择领域,效应较大且不同遗传背景下稳定的分子标记,大多数与植物抗逆性相关。
其原因笔者认为主要有两点:
第一,抗逆相关性状需要在特定的环境条件下才能表现出来,因此在人类长期以来对作物的驯化过程中,受到的选择压力较小。换句话说,就是依然保留在少数材料中的抗性位点相对比较容易发现。尤其是产量较低的地方种、祖先种及近缘物种,往往抗逆性较强,存在可以挖掘利用的抗性相关基因。
第二,与株型、产量等性状相比,某种针对性的抗性往往只牵扯到少数激素、代谢物或蛋白的合成,因此参与调控的基因较少,也就更容易找到效应较大的主效基因或位点。
理所当然的,在基因编辑作物改良领域,抗逆性改良也出现了大量的成功案例。
做玉米育种的同仁应该了解,穗粒腐病是世界范围内的一种重要病害,在我国自东北到西南均发病严重。现在国家的玉米审定标准中,对穗粒腐病高感的品种实行一票否决制。然而高抗穗粒腐病的材料很少,在性状的鉴定及改良上也存在巨大困难。2022年8月,谢传晓团队发表在PBJ上的论文,证实他们通过基因编辑敲除玉米第三染色体上ZmFER1(小麦赤霉病抗性相关基因Fhb1的同源基因),从而提升了玉米对镰孢菌穗腐病的抗性。与改造前的材料相比,野生型中伏马毒素的含量下降到了千分之一以下。更为关键的是,改造后的材料在其他农艺性状上未发现显著的差异(Liu et al., 2022)!
在麦类作物上,康振生院士团队利用CRISPR/Cas9基因编辑技术敲除小麦TaCIPK14基因,使小麦产生了对5个条锈菌小种的广谱抗性,同时没有发现显著的农艺性状差异(He et al., 2023)。通过敲除PDIL5-1基因创造的大麦抗花叶病种质,也被证明对植株的生长和产量没有影响(Hoffie et al., 2023)。2023年1月30日,PBJ上的另一篇研究论文,科学家利用基因组编辑方法创制番茄eIF4E1基因的非同义突变,赋予了番茄对多种病毒的抗性。
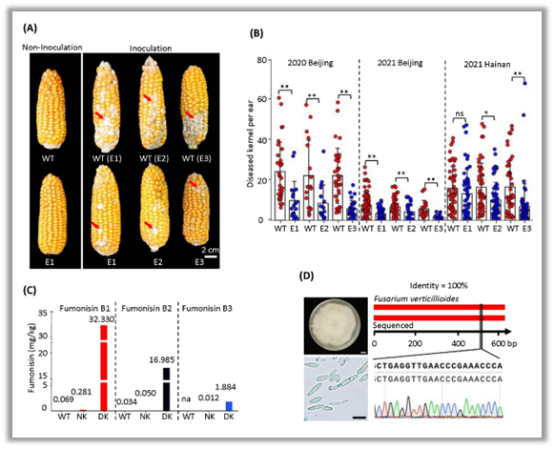
基因编辑敲除ZmFER1提高玉米对穗粒腐病的抗性(Liu et al., 2022)
在非生物胁迫抗性改良方面,陕西省杂交油菜研究中心张彦锋团队通过基因编辑靶向敲除核孔蛋白复合体外环结构成员之一的BnHOS1,成功获得了油菜抗寒新种质(Song et al., 2022)。在抗虫性改良方面,安徽农业大学李培金团队发现基因编辑突变RPX1基因后植物对小菜蛾的抗性提高,更为重要的是对rpx1的植物种子产量没有显著负面影响(Chen et al., 2022)。
除了上述抗病、抗虫、抗非生物胁迫的基因编辑改良案例外,还有大量基因编辑提升植物抗逆性的相关报道。尤其是在以往的研究中,已经有大量负调控抗性的作物基因被克隆,这些基因都可以作为潜在基因编辑靶点用以创制抗性增强的植物品种。
然而需要注意的是,在许多研究中并未检测基因编辑对植物其他性状是否发生显著影响。鉴于许多抗性相关基因曾被报道会影响抗性及植物生长的平衡关系,即便针对一些功能明确的抗性相关基因进行基因编辑,仍需考虑大田验证来确保其对其他生产性状不会造成大的影响。
二、作物品质改良
20世纪之前,作物生产的主要需求都是解决粮食紧缺的问题,因此产量是育种家们最为关注的性状。到了21世纪,我国居民的生活水平飞速提升,老百姓对于食物品质的追求愈发受到关注。
由于很多品质性状在过去很难通过肉眼进行选择,因此在这类性状上具有较丰富的遗传变异。随着大量品质相关基因的克隆,基因编辑在此方向的应用日渐广阔。
在这之中,以油料作物的基因编辑改良探索最为广泛。以油菜为例,提升胡萝卜素、提升维生素E含量、降低葡萄糖苷含量、增加含油量、提升油酸含量、降低芥酸含量等品质性状的基因编辑案例近年来均有文章报道,且大部分已经递交了国家发明专利申请。在花生上,张新友院士也谈到我国已利用基因编辑技术创制出高油酸的花生品种,其油酸含量可以超过75%。在大豆上,最近的一篇报道显示,福建农林大学/广州大学关跃峰团队利用CRISPR/Cas9多基因编辑技术创制了几乎不含低聚糖的大豆新种质,进而可以避免肠道微生物发酵造成胀气,同时可以提高牲畜饲料的能量利用效率(Lin et al., 2023)。
减少有害物质的积累也是作物品质改良的一个重要方向。木薯是非洲许多地区的主粮,然而鲜木薯中含有较高的生氰糖苷类物质亚麻苦甙,生氰糖苷降解后产生的有毒氢氰酸会给食用人群造成风险。科学家通过基因编辑敲除木薯MeCYP79D1基因,获得了氰化物含量降低的基因编辑木薯材料,其株系中的亚麻苦甙和氰化物含量降低了7倍多(Juma et al., 2022)。
在主粮作物上,关于糯性及甜性相关的主要基因早已被克隆,与之相应的通过基因编辑快速改良相关性状的专利早已被大量申请。此外,提高类胡萝卜素含量、降低醇溶蛋白含量等方面,基因编辑也已经获得了成功。一项有趣的研究是,科学家根据香稻香气来源物质2‐acetyl‐1‐pyrroline的积累原理,利用基因编辑技术创造了带有香味的玉米和高粱品种(Wang et al., 2021)。最近,中国农业科学院农业基因组研究所张翠军团队通过CRISPR/Cas9基因编辑技术,成功将白粒小麦转化为富含花青素的红粒小麦,并提高了小麦的抗穗发芽能力(Zhu et al., 2022)。
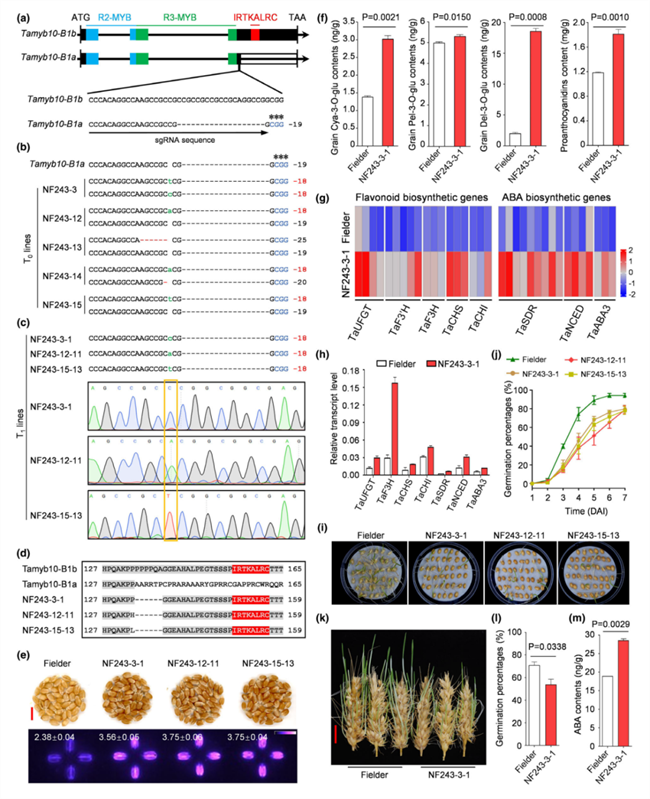
基因编辑创制的红粒小麦穗发芽能力提升(Zhu et al., 2022)
三、产量及株型改良
相较于抗逆性及品质性状来说,主要粮食作物可以发掘的新增产基因较少,这主要是由于大多数主效的产量相关基因已经在数千年的育种进程中受到了选择。
令人欣喜的是,在此基础上仍然有一些通过基因编辑增加作物产量的报道。
其中,最令人瞩目的是中国农业大学杨小红团队在Science上发表的文章。他们发现玉米和水稻的数量性状位点KRN2经历了趋同选择,且负调控两种作物的粒数。通过基因编辑在玉米和水稻中敲除KRN2,籽粒产量分别提高了10%和8%,而其他农艺性状无明显损失(Chen et al., 2022)。就玉米而言,与对照相比产量提升5%就可以达到新品种审定的标准,10%的产量提升意义可见一斑。
在另一种主粮小麦上,美国南达科他州立大学通过基因编辑改变TaSPL13基因末端的microRNA156识别元件,从而令小麦开花提前、分蘖数和株高减少、粒宽和粒数增加。一个基因的编辑同时造成了多个产量和株型性状的改善,揭示了TaSPL13基因在提高小麦产量方面具有巨大的潜力(Gupta et al., 2022)。
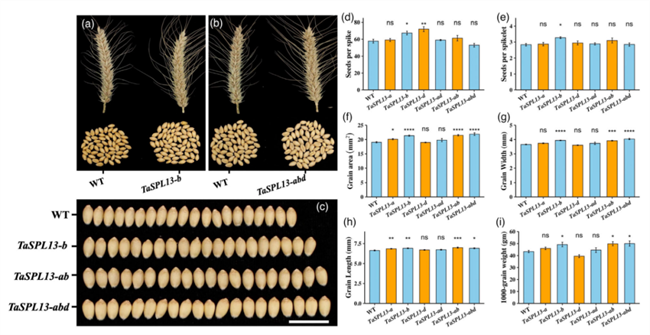
基因编辑TaSPL13基因可以提高小麦产量(Gupta et al., 2022)
在株型方面,20世纪第一次绿色革命后人们开始认识到作物矮化在生产上的巨大优势,各类相关基因也逐渐被发掘和利用,至今仍有一些矮化基因和株型紧凑基因在不断克隆和报道。2022年12月,来自中国农业科学院蔬菜花卉研究所、深圳农业基因组所、西北农林科技大学、北京市农林科学院蔬菜研究所等4家单位合作,在一个南瓜稀有矮化变异基因的基础上,通过基因编辑技术针对性地在其他多种瓜类作物中进行基因变异,创造出原本自然界不存在的紧凑株型材料,大大提高了葫芦科瓜类作物的生产效率,显著节省了劳动力(Wang et al., 2022)。
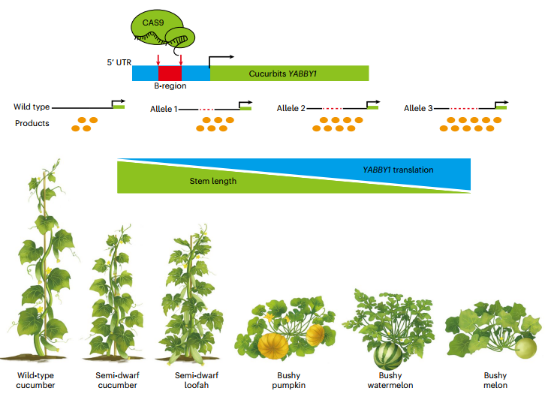
基因编辑创制株型紧凑葫芦科植物(Wang et al., 2022)
其他关于产量和株型基因编辑的报道还有一些,但由于大多数产量及株型相关性状都是微效多基因控制的数量性状,效应较大的主效基因较少。且在驯化时间较长的主要作物上,大多数主效位点的有利变异已经被我们的祖先选择利用。因此,大多数克隆的产量及株型相关基因对表型的变异贡献率都相对较小。
那么,利用杂种优势且遗传转化受基因型限制的作物(如玉米),需要特别注意。如果待改良的品种不能够直接进行遗传转化,需要回交导入基因编辑创造的基因型的话,此时引入小效应的增产基因型的作用可能无法抵消回交导致的杂种优势下降。
因此,利用基因编辑进行主要作物的产量改良,需要有更严谨的试验设计和接受非预期结果的心理准备。
四、特殊性状的创造
与产量性状不同,许多非植物生活必需的特殊性状,往往本身就是基因异常突变产生的。对这类性状,基因编辑可以对目标作物进行快速的改造。
在已经成功的大量例子中,比较常见的包括:雄性不育系创制、除草剂抗性创制、单倍体诱导系创制、解除遗传转化的基因型限制,以及其他一些新的用途。
通过基因编辑创造雄性不育系的报道和专利很多,然而往往没有对应的保持系创制手段,因此实际使用具有一定的难度,在这里笔者不展开讨论。
对于其他几个方面,会依次进行举例说明。
作物除草剂抗性的产生常归于三种机理:1)除草剂靶标蛋白的高表达弥补了除草剂的伤害;2)除草剂靶标蛋白发生变异从而降低了与除草剂的结合能力;3)通过特定途径快速对除草剂进行降解。现有的抗除草剂转基因品种,就是通过转入细菌的关键酶降解除草剂,即第三种抗除草剂机理。而利用基因编辑创制抗除草剂基因,往往是利用第二种机理,即通过基因编辑使除草剂的靶蛋白产生变异,在保持其生物功能的基础上降低其与除草剂的结合能力。
通过这种原理,我国科学家在水稻、小麦、大豆、花生、油菜、西瓜等多种作物上成功创造出了针对不同除草剂的抗性基因。最近,中国农业科学院植物保护研究所与农业基因组研究所合作,对水稻内源基因OsGS1进行碱基编辑,获得了3种抗广谱除草剂草铵膦的等位基因。载药平板萌发和草铵膦(田间推荐剂量)喷施试验结果均证实,携带这些等位基因的水稻具有抗草铵膦的能力(Ren et al., 2022)。
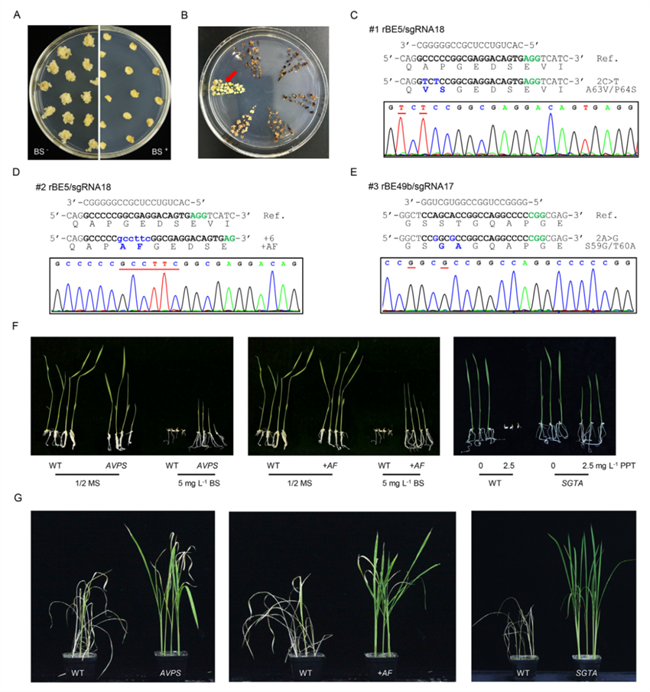
利用基因编辑创制抗除草剂水稻(Ren et al., 2022)
单倍体诱导系因为可以用来快速创制纯合株系,所以在育种上具有非常重要的价值。目前最成熟的商用单倍体诱导技术来自于玉米Stork6衍生系,可以通过给目标材料授粉诱导产生母本单倍体。现在的研究表明其诱导能力主要来自于ZmPLA1(MTL/NLD)基因的突变,同时ZmDMP基因的突变也在诱导率的提升上具有一定的贡献。近些年的研究发现,基因ZmPLD3突变可以产生或提升玉米的单倍体诱导能力。这3个基因的突变都已经被证实可以通过基因编辑简单的敲除来创制,且可以推广到其他单子叶植物。去年11月,中国农业科学院作物科学研究所叶兴国团队通过基因编辑HvMTL基因,创制了大麦单倍体诱导系,并且达到了10%-12%的诱导率(Tang et al., 2022)。
在双子叶植物上,未发现ZmPLA1介导的单倍体诱导机制。但是编辑改造着丝粒组蛋白酶3(CENH3),可以创造出父本或母本的单倍体诱导系。
许多植物的愈伤组织再生能力过低,限制了它们遗传改良的进程。为了克服基因型对遗传转化的限制,科学家们做了许多的工作。最近的研究显示,利用基因编辑技术突变苹果MdSPL6基因,可以显著提高其不定芽的再生效率,为在苹果和其他蔷薇科果树中创制高再生种质揭示了一种新的可行方案(Li et al., 2022)。
近期,能够让作物种子始终保持杂种优势的作物无融合生殖技术,也依靠基因编辑取得了重大突破。Nature Communications上的一篇文章显示,将CRISPR/Cas9介导的MiMe失活系统和孤雌生殖诱导系统一次性构建到同一个T-DNA上,在卵特异性启动子的作用下,可以在水稻的F₁杂种中实现高效无融合生殖,二倍体比率大大提高(90%以上),且无融合生殖植物在连续世代中保持F₁杂种的表型。尽管该技术由于各种原因距离商业应用尚早,但这也显示出了基因编辑技术用来开发功能性作物种质资源的巨大潜力(Vernet et al., 2022)。
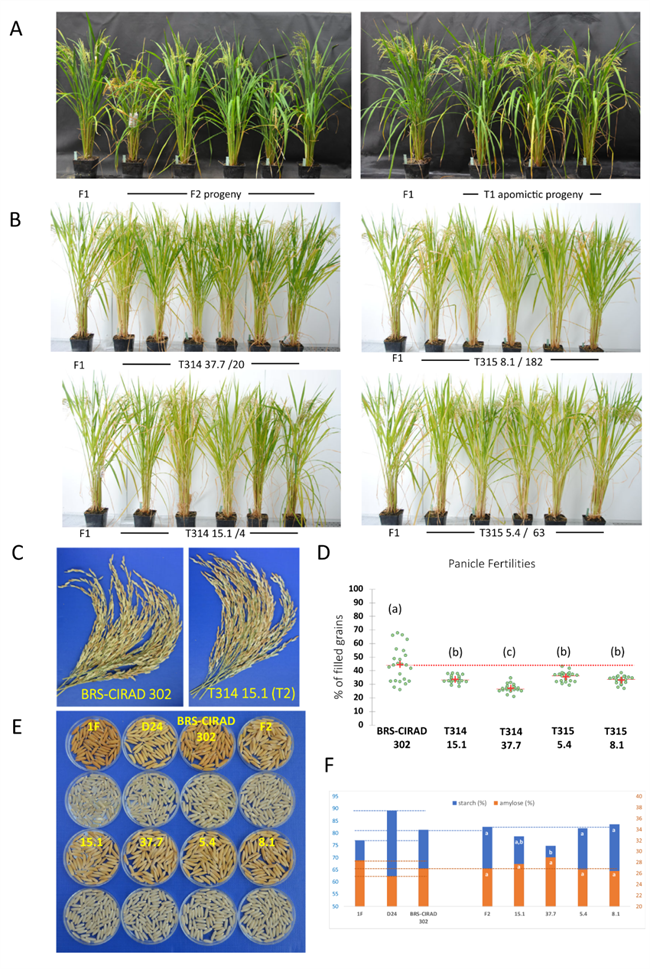
基因编辑创制无融合生殖水稻(Vernet et al., 2022)
本次分享的内容较多,感谢耐心看到这里的各位同仁。
按照一开始我们提出的问题,下一期的文章将对基因编辑品种创制的方法、流程及需要消耗的金钱及时间成本进行分享和讨论。敬请关注下期文章《新时代对基因编辑作物商业化的展望与思考(下)》。
参考文献
Chen H, Chen C, Huang S, Zhao M, Wang T, Jiang T, Wang C, et al. Inactivation of RPX1 in Arabidopsis confers resistance to Plutella xylostella through the accumulation of the homoterpene DMNT. Plant, Cell & Environment, 2022, doi: 10.1111/pce.14528.
Chen W, Chen L, Zhang X, Yang N, Guo J, Wang M, Ji S, et al. Convergent selection of a WD40 protein that enhances grain yield in maize and rice. Science, 2022, 375(6587): eabg7985.
Gupta A, Hua L, Zhang Z, Yang B, Li Wang. CRISPR-induced miRNA156-recognition element mutations in TaSPL13 improve multiple agronomic traits in wheat. Plant Biotechnology Journal, 2022, doi: 10.1111/pbi.13969.
He F, Wang C, Sun H, Tian S, Zhao G, Liu C, Wan C, et al. Simultaneous editing of three homoeologs of TaCIPK14 confers broad-spectrum resistance to stripe rust in wheat. Plant Biotechnology Journal, 2023, 21(2): 354–368.
Hoffie RE, Perovic D, Habekuß A, Ordon F, Kumlehn J. Novel resistance to the Bymovirus BaMMV established by targeted mutagenesis of the PDIL5-1 susceptibility gene in barley. Plant Biotechnology Journal, 2023, 21(2): 331–341.
Huali Tang, Yuliang Qiu, Wanxin Wang, Mei Yu, Yannan Chang, Zhiyang Han, Lipu Du, Zhishan Lin, Ke Wang, Xingguo Ye. (2022). Development of a haploid-inducer by editing HvMTL in barley. Journal of Genetics and Genomics.
Juma BS, Mukami A, Mweu C, Ngugi MP, Mbinda W. Targeted mutagenesis of the CYP79D1 gene via CRISPR/Cas9-mediated genome editing results in lower levels of cyanide in cassava. Frontiers in Plant Science, 2022, 13: 1009860.
Li H, Sun H, Dong H, Wang S, Fan X, Li Y, Cheng L, et al. Genome editing of apple SQUAMOSA PROMOTER BINDNG PROTEIN-LIKE 6 enhances adventitious shoot regeneration. Plant Physiology, doi:10.1093/plphys/kiac570.
Lin W, Kuang H, Bai M, Jiang X, Zhou P, Li Y, Chen Bo, et al. Multiplex genome editing targeting soybean with ultra-low anti-nutritive oligosaccharides. The Crop Journal, 2023, doi: 10.1016/j.cj.2023.01.001.
Liu C, Kong M, Zhu J, Qi X, Duan C, Xie C. Engineering null mutants in ZmFER1 confers resistance toear rot caused by Fusarium verticillioidesin maize. Plant Biotechnology Journal, 2022, 20(11): 2045–2047.
Ren B, Kuang Y, Xu Z, Wu X, Zhang D, Yan F, Li X, et al. Three novel alleles of OsGS1 developed by base-editing-mediated artificial evolution confer glufosinate tolerance in rice. The Crop Journal, 2022, doi: 10.1016/j.cj.2022.10.003.
Song M, Bin L, Huang S, Hu S, An R, Wei S, Mu J, Zhang Y. Identification of nuclear pore complexes (NPCs) and revealed outer-ring component BnHOS1 related to cold tolerance in B. napus. International Journal of Biological Macromolecules, 2022, 223: 1450-1461.
Vernet A, Meynard D, Lian Q, Mieulet D, Gibert O, Bissah M, Rivallan R, et al. High-frequency synthetic apomixis in hybrid rice. Nature Communication, 2022, 13(1):7963.
Wang S, Wang K, Li Z, Li Y, He J, Li H, Wang B, et al. Architecture design of cucurbit crops for enhanced productivity by a natural allele. Nature Plants, 2022, 8(12): 1394-1407.
Wang Y, Liu X, Zheng X, Wang W, Yin X, Liu H, Ma C, et al. Creation of aromatic maize by CRISPR/CasFA. Journal of Integrative Plant Biology, 2021, 63(9): 1644-1670.
Zhu Y, Lin Y, Fan Y, Wang Y, Li P, Xiong J, He Y, et al. CRISPR/Cas9-mediated restoration of Tamyb10 to create pre-harvest sprouting-resistant red wheat. Plant Biotechnology Journal, 2022, doi: 10.1111/pbi.13981.






